and clinical posturology
Pierre-Marie GAGEY
Institut de Posturologie, Paris
Lecture at the Collège de France in Paris on Saturday the 16th of November, 1996
to the members of the Association Française de Strabologie interested in Posture.
The
perception of space is not built independently on each sensitivo-sensory
canal. This very old hypothesis, that Aristotle already proposed
in his Peri Psuches, has been studied for the perception
of visual space that is built from retinal afferences and proprioceptive
oculomotor afferences (Blakemore et al., 1975; Cynader
et al., 1975; Cynader et al., 1976; Hein et al.,
1979; Hein et al., 1987).
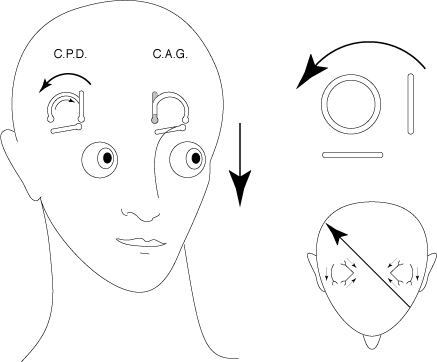
In order to check that this hypothesis is also true concerning visual and vestibular afferences, we could take into account the very particular anatomy and the physiology of the semi-circular canals. Each canal, indeed, specifically perceives angular accelerations depending on the plane, the way and the direction of the acceleration relatively to the plane of that canal (Ross, 1936; Löwenstein & Sand, 1940; Adrian, 1943; Zotterman, 1943; Estes et al., 1975; Banks et al., 1975.a). Thus, an angular acceleration parallel to the plane of the right posterior canal and going backwards and right will involve the appearance of stimulating afferences only from the cupula of that canal (fig. 1). Now, in the absence of movements of the eyeballs, the head movements provoked by those angular accelerations are accompanied on the retina by a shift of the environment image along an ocular meridian parallel to the vector of that angular acceleration. If, during the ontogenesis, the perception of space is not built independently for the visual canal and for the vestibular canal, signs of this privileged relation between the planes of the semi-circular canals and the directions of the visual space have to be found.
FIG. 1 - An angular acceleration in the plane of the right posterior canal (CPD) involves a movement of the stimulating endolymph only in the right posterior canal. If the eyeball is motionless, the movement induced by this acceleration is accompanied by a retinal shift along an ocular meridian parallel to the plane of the right posterior canal, in the way and direction of the acceleration.
Approximately knowing the planes of the semi-circular canals, it is possible to determine geometrically those ocular meridians «corresponding» to the planes of the semi-circular canals. The plane of the left posterior canal, for instance, makes an angle of around 71° relatively to the horizontal plane of the stereotaxic system of Reid (q1 on figure 2) and an angle of around 55° relatively to the sagittal plane of the same system (q2 on figure 2) (Blanks et al., 1975.b). The section of the eyeball by a plane parallel to the plane of that canal and going through the centre of the eyeball, corresponds to the meridian along which the environment image shifts during movements with accelerations stimulating the left posterior canal.
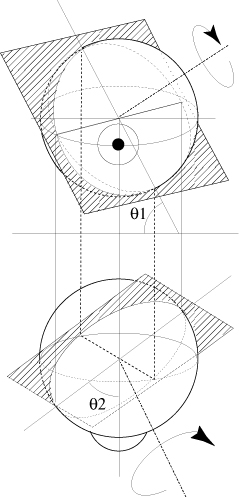
FIG. 2 - Descriptive
study of the section of an eyeball by a plane parallel to the
plane of the left posterior canal and going through the centre
of the eye.
q1= 55°, q2 = 71° relatively to the
stereotaxic planes of Reid (Blanks et al., 1975.b)
That last remark points to a privileged relation between each semi-circular canal and one or several oculomotor muscles that, when close to the primary position, provoke a rotation around an axis perpendicular to their plane. That privileged relation has actually already been described by various authors (Lorente de No, 1932; Szentagothai, 1950; Cohen et al., 1964; Ito et al., 1976).
hypothesis
. A
prism deviates the visual space in a single direction that can
be identified thanks to the position of the base of the prism.
If, during the ontogenesis, the perception of space has been built
coherently for the visual and the vestibular sensory canals, certain
relation between the planes of the canals and the position
of the base of the prism has to be found. For instance, a
prism with a base situated at 125° provokes on the retina
a deviation of the visual space approximately parallel to the
retinal meridian «associated» to the plane of the
right posterior canal (fig. 3).
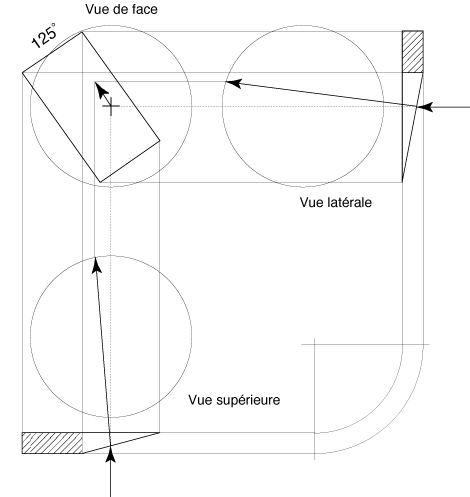
FIG. 3 - Descriptive
study of the deviation of the visual space by a prism with a
base situated at 125°.
Vue de face = Front view,
Vue latérale = lateral view,
Vue supérieure = superior view
The deviation of the visual space is approximately parallel to
the retinal meridian "associated" to the right posterior
semi-circular canal.
. The apparent movement of the environment image on the
retina of the right eye corresponds to the movement of the eyeball,
close to the primary position, produced by the action of the obliquus
inferior muscle.
. In order
to check that this relation between the planes of the canals
and the position of the base of the prism can be found again
experimentally, a retrospective study has been undertaken on the
files of postural patients of the Institut de Posturologie treated
by the wearing of optical prisms.
. Since the publications of Magnus (1924), we know, indeed,
that the perception of space intervenes in the regulation of the
tonus of posture. It is therefore possible to propose a study,
via postural tonus, of phenomena intervening in the perception
of space.
Subjects
. The
files of 197 patients who had been attended at the Institut de
Posturologie for symptoms related to functional disorders of the
control of the orthostatic posture, have been selected because
they contained precise reports of normalised postural clinical
examinations (Gagey, 1993) all those patients had undergone. Those
files notably contained numbered evaluations of postural tonus,
with and without an optical prism of which the position of the
base was known.
. Those patients
either complained of vertigo sensations and/or instability, without
anomalies in the vestibular functional tests, or of various pains
in the corporal axis in orthostatism (lumbar pains, cervical pains,
etc.).
Evaluation of postural tonus
. The
numbered evaluation of postural tonus selected for this study
is the spin movement observed during the stepping test of Fukuda-Unterberger
(Fukuda, 1959). That spin parameter has been studied in a population
of normal subjects; we know the distribution of its random fluctuations
between two tests performed in similar conditions at a few minutes
or a few weeks interval: the mean of matching differences between
two similar tests is of 0° ± 25° (Weber et al.,
1984). The performance conditions of the stepping test described
by Gagey & Weber (1995) have been respected in the population
of patients studied.
We know that changes in the symmetry of postural tonus involve
systematic changes in the spin movement during the stepping test
(Fukuda, 1959; Ushio et al., 1976).
Position of the base of the prism
. In
accordance with what was demonstrated in the introduction, the
studied positions of the base of the prism are - written in the
trigonometrical order from a left temporal [or right nasal] position:
0°, 55°, 125°, 180°, 235°, 305°.
. All the
subjects of this study have performed a stepping test without
prism and a stepping test with prism in at least one of those
positions of the base of the prism. No subject has repeated the
stepping test for all the positions of the base of the prism.
The position of the base of the prism studied in a given subject
has been determined by the other tests of his postural clinical
examination that showed the possibility to modify the postural
tonus of that subject thanks to a prism of three prismatic dioptres
put in that position.
. The distribution of the changes in the spin movement
provoked by the wearing of the prism is very significantly different
from the distribution of the random changes of that same movement
(p<0.001), whatever the position of the base of the prism (Fig.
4).
. The way
the spin movement changes depends on the position of the base
of the prism (fig. 4): sometimes, under the effect of the prism,
the subject turns more to his right when the base of the prism
is put at 0° or 125° or 235° (negative values of the
spins difference with and without prism), sometimes he turns more
to his left when the base of the prism is put at 55° or 180°
or 305° (positive values of the spins difference with and
without prism).
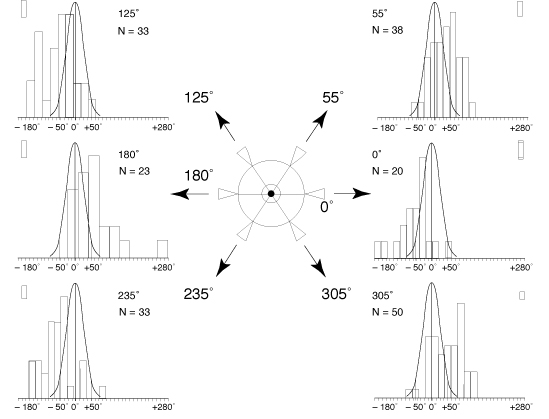
FIG. 4 - Spin changes
observed according to the different positions of the base of
the prism.
At the centre: diagram showing the different studied positions
of the base of the prism put before the patient's eye (either
left or right).
On the sides, for each of the six positions of the base of the
prism, histogram of the change in the spin movement provoked
by the placing of the prism. Curve of Gauss reminding the normal
theoretical distribution of the random changes of the spin movement.
Discussion
. The bearing of this statistical analysis is limited.
It only concerns subjects who showed functional disorders related
to the orthostatic posture. However, a similar study has been
carried out on normal subjects (Ushio et al., 1980), but
it only concerned positions 0° and 180° of the base of
the prism and it was done on a small number of subjects only.
Yet the results of the study of Ushio and colleagues agree with
the present analysis.
. Each subject
has only been studied in one position of the base of the prism;
the described differences between all the positions of the base
of the prism could therefore be seen as interindividual differences.
However a study, close to the works presented in this article,
has been done on normal subjects with turning prisms that, in
eight seconds, explored in the same subject all possible positions
of the base of the prism (Séverac et al., 1993).
Now, the results of Séverac and colleagues also show a
difference of direction of the postural answer according to the
position of the base of the prism. The agreement of the results
of Séverac and colleagues with the present study ends here
because the differences between the protocols do not allow a more
precise conclusion.
. The reported clinical facts do not allow proving that
the perception of space, during the ontogenesis, is organised
in a coherent way between the visual and vestibular sensory canals,
yet they certainly make that hypothesis even more probable.
References
Adrian E.D. (1943) Discharges from vestibular receptors
in the cat. J. Physiol. (London), 101, 389-407.
Blakemore C, Van Sluyters RC, Peck CK, Hein A (1975) Development
of cat visual cortex following rotation of one eye. Nature,
257, 584-6
Blanks R.H.I., Estes M.S., Markham C.H. (1975.a) Physiologic caracteristics
of vestibular first-order canal neurons in Cat. II. Response to
constant angular acceleration. J. Neurophysiol., 38,
1250-68.
Blanks R.H.I., Curthoys I.S., Markam C.H. (1975.b) Planar relationships
of the semicircular canals in man. Acta Otolaryngol. (Stockh.),
80, 191-200.
Cohen B., Suzuki J., Shanzer S., Bender M.B. (1964) Semi-circular
control of eye movements. In: M.B. Bender (Ed.) The oculomotor
system. Harper & Row, New York.
Cynader M, Berman N, Hein A (1975) Cats raised in a one-directional
world: effects on receptive fields in visual cortex and superior
colliculus. Exp Brain Res, 22, 267-80
Cynader M, Berman N, Hein A (1976) Recovery of function in cat
visual cortex following prolonged deprivation. Exp Brain Res,
25, 139-56
Estes M.S., Blanks R.H.I., Markham C.H. (1975) Physiologic caracteristics
of vestibular first-order canal neurons in Cat. I. Response plane
determination and resting discharge characteristics. J. Neurophysiol.,
38, 1232-49.
Fukuda T. (1959) The stepping test. Two phases of the labyrinthine
reflex. Acta Otolaryngol. (Stockh.) 50, 2: 95-108.
Gagey P.M. (1993) Le bilan postural. Ann. Kinésithér.,
20, 295-301.
Gagey P.M., Weber B. (1995) Posturologie; Régulation
et dérèglements de la station debout. Masson,
Paris.
Hein A (1987) La structuration de l'espace visuel peut-elle se
développer en l'absence des informations proprioceptives
oculomotrices? Critique de la Posturologie, 12,
1-4.
Hein A, Vital-Durand F, Salinger W, Diamond R (1979) Eye movements
initiate visual-motor development in the cat. Science,
204, 1321-2
Ito M., Nisimaru N., Yamamoto M. (1976) Pathways for the vestibulo-ocular
reflex excitation arising from semicircular canals of rabbits.
Exp. Brain Res., 24, 257-271.
Lorente de Nò R. (1932) The regulation of eye positions
and movements induced by labyrinth. Laryngoscope, 42,
233-332.
Löwenstein O. & Sand A. (1940) The individual and integrated
activity of the semicircular canals of the elasmobranch labyrinth.
J. Physiol. (London), 99, 89-101.
Magnus R. (1924) Körperstellung. Springer (Berlin).
Ross D.A. (1936) Electrical studies on the frog's labyrinth.
J. Physiol. (London), 86, 117-46.
Séverac A., Bessou P., Pagès B. (1993) Unusual visual
stimulation in dynamic balance conditions: proposal for a space
motion sickness test. Advances in space research,
Szentágothaì J. (1950) The elementary vestibulo-ocular
reflex arc. J. Neurophysiol., 13, 395-407.
Ushio N., Hinoki M., Baron J.B., Gagey P.M., Meyer J. (1976) The
stepping test with neck torsion: proposal of a new equilibrium
test for cervical vertigo. Practica Otologica Kyoto, 69,
Sup.3, 1369-79 (En japonais).
Ushio N., Hinoki M., Nakanishi K., Baron J.B. (1980) Rôle of oculomotor proprioception in the maintenance of body equilibrium; correlation with the cervical one. Agressologie, 21,
E, 143-52.
Weber B., Gagey P.M., Noto R. (1984) La répétition
de l'épreuve modifie-t'elle l'exécution du test
de Fukuda? Agressologie, 25, 1311-14.
Wolgin DL, Hein A, Teitelbaum P (1980) Recovery of forelimb placing
after lateral hypothalamic lesions in the cat: parallels and contrasts
with development. J Comp Physiol Psychol, 94, 795-807
Zotterman Y. (1943) The microphonic effect of teleost labyrinth
and its biological significance. J. Physiol. (London),
102, 313-8.