Loi des canaux
et clinique posturale
Pierre-Marie GAGEY
Institut de Posturologie, Paris
Conférence donnée au Collège de France, Paris, le samedi 16 novembre 1996
aux membres de l´Association Française de Strabologie interessés par la Posture
Compte-rendu par P.V. Bérard.
Introduction
La perception de l´espace
ne se construit pas indépendamment sur chaque canal sensitivo-sensoriel.
Cette hypothèse très ancienne, Aristote la proposait
déjà dans son peri Psuches, a été
étudiée pour la perception de l´espace visuel
qui se construit à partir des afférences rétiniennes
et des afférences proprioceptives oculomotrices (Blakemore
et al., 1975; Cynader et al., 1975; Cynader et al.,
1976; Hein et al., 1979; Hein, 1987). Pour vérifier
que cette hypothèse est aussi vraie en ce qui concerne
les afférences visuelles et vestibulaires, il est possible
de tenir compte de l´anatomie très particulière
et de la physiologie des canaux semi-circulaires. Chaque canal,
en effet, perçoit d´une manière spécifique
les accélérations angulaires selon le plan, le sens
et la direction de l´accélération par rapport
au plan de ce canal (Ross, 1936; Loewenstein & Sand, 1940;
Adrian, 1943; Zotterman, 1943; Estes et al., 1975; Blanks
et al., 1975.a). Ainsi une accélération angulaire
parallèle au plan du canal postérieur droit et dirigée
en arrière et à droite entraînera l´apparition
d´afférences excitatrices uniquement à partir
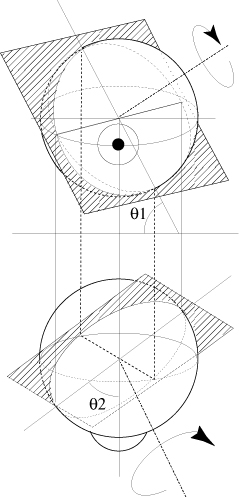
de la cupule de ce canal (fig. 1).
Or,
en l´absence de mouvements des globes oculaires, les mouvements
de la tête provoqués par ces accélérations
angulaires s´accompagnent sur la rétine d´un
glissement de l´image de l´environnement, qui se produit
le long d´un méridien oculaire parallèle au
vecteur de cette accélération angulaire. Si, au
cours de l´ontogenèse, la perception de l´espace
ne se construit pas indépendamment pour le canal visuel
et pour le canal vestibulaire, on doit retrouver des traces de
cette liaison privilégiée entre les plans des canaux
semi-circulaires et les directions de l´espace visuel.
FIG. 1 - Une accélération
angulaire dans le plan du canal postérieur droit provoque
un mouvement de l´endolymphe excitateur uniquement dans
le canal postérieur droit. Si le globe oculaire est immobile,
le mouvement induit par cette accélération s´accompagne
d´un glissement rétinien le long d´un méridien
oculaire parallèle au plan du canal postérieur droit,
dans le sens et la direction de l´accélération.
Connaissant approximativement
les plans des canaux semi-circulaires il est possible de déterminer
géométriquement ces méridiens oculaires qui
«correspondent» aux plans des canaux semi-circulaires.
Le plan du canal postérieur gauche, par exemple, fait un
angle de 71° environ par rapport au plan horizontal du système
stéréotaxique de Reid (Tétha1 sur la figure
2) et un angle de 55° environ par rapport au plan sagittal
du même système (Tétha2 sur la figure 2) (Blanks
et al., 1975.b). La section du globe oculaire par un plan parallèle
au plan de ce canal et passant par le centre du globe correspond
au méridien le long du quel glisse l´image de l´environnement
lors des mouvements dont les accélérations stimulent
le canal postérieur gauche.
On
peut remarquer que le même glissement rétinien est
provoqué par une rotation du globe oculaire autour d´un
axe perpendiculaire à ce plan.
FIG. 2 - Étude descriptive de la
section d´un globe oculaire par un plan parallèle
au plan du canal postérieur gauche et passant par le centre
de l´oeil. Tétha1 = 55°, tétha2 = 71°
par rapport aux plans stéréotaxiques de Reid (Blanks
et al., 1975.b)
Cette dernière remarque
permet de prévoir une relation privilégiée
entre chaque canal semi-circulaire et le ou les muscles oculomoteurs
qui, au voisinage de la position primaire, provoquent une rotation
autour d´un axe perpendiculaire à leur plan. Cette
relation privilégiée a, de fait, été
décrite par plusieurs auteurs (Lorente de Nò, 1932;
Szentágothaì, 1950; Cohen et al.,1964; Ito
et al.,1976).
Un
prisme dévie l´espace visuel dans une seule direction
qu´on peut repérer par la position de la base de
prisme. Si, au cours de l´ontogenèse, la perception
de l´espace s´est construite de manière cohérente
pour les canaux sensoriels visuel et vestibulaire, on doit retrouver
une certaine relation entre les plans des canaux et la position
de la base du prisme. Un prisme, par exemple, dont la base
est située à 125° provoque sur la rétine
une déviation de l´espace visuel approximativement
parallèle au méridien rétinien «associé»
au plan du canal postérieur droit (fig. 3).
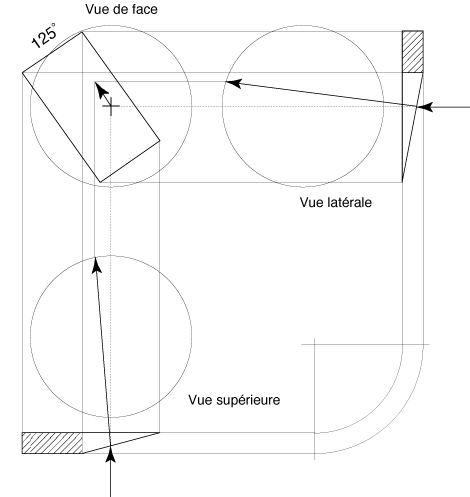
FIG. 3 - Étude descriptive de la
déviation de l´espace visuel par un prisme dont la
base est située à 125°.
La déviation de l´espace
visuel est approximativement parallèle au méridien
rétinien «associé» au canal semi-circulaire
postérieur droit.
Le mouvement apparent de
l´image de l´environnement sur la rétine de
l´oeil droit correspond à un mouvement du globe oculaire,
au voisinage de la position primaire, produit par l´action
du muscle petit oblique.
Pour vérifier que
cette relation entre les plans des canaux et la position de
la base du prisme peut être retrouvée expérimentalement,
une étude rétrospective a été entreprise
sur les dossiers de malades posturaux de l´Institut de Posturologie
traités par le port de prismes optiques.
Depuis les publications de
Magnus (1924) on sait, en effet, que la perception de l´espace
intervient dans la régulation du tonus de posture. Il est
donc possible de proposer une étude, par le biais du tonus
postural, de phénomènes intervenant dans la perception
de l´espace.
Matériel et méthodes
Sujets
Les dossiers de 197 patients,
qui avaient consulté à l´Institut de Posturologie
pour des symptômes en rapport avec des troubles fonctionnels
du contrôle de la posture orthostatique, ont été
sélectionnés parce qu´ils contenaient un compte-rendu
précis d´un examen clinique postural normalisé
(Gagey, 1993) auquel tous ces patients avaient été
soumis. Ces dossiers comportaient en particulier une évaluation
chiffrée de tonus postural, avec et sans port d´un
prisme optique dont la position de la base était connue.
Ces
malades se plaignaient soit de sensations vertigineuses et/ou
d´instabilité, sans anomalies aux épreuves
fonctionnelles vestibulaires, soit d´algies diverses de
l´axe corporel en orthostatisme (lombalgies, cervicalgies,
etc.).
Évaluation du tonus postural
L´évaluation
chiffrée du tonus postural retenue pour cette étude
est le mouvement de spin observé au cours du test de piétinement
de Fukuda-Unterberger (Fukuda, 1959). Ce paramètre de spin
a été étudié dans une population de
sujets normaux, on connaît la distribution de ses fluctuations
aléatoires entre deux tests pratiqués dans des conditions
semblables à quelques minutes ou quelques semaines d´intervalle:
la moyenne des différences appariées entre deux
tests semblables est de 0° ± 25° (Weber, et
al., 1984). Les conditions d´exécution du test
de piétinement décrites par Gagey & Weber (1995)
ont été respectées dans la population de
malades étudiés.
On
sait que des variations de la symétrie du tonus postural
induisent des variations systématiques du mouvement de
spin au cours du test de piétinement (Fukuda, 1959; Ushio
et al., 1976).
Positions de la base du prisme
Conformément à
ce qui a été exposé dans l´introduction,
les positions étudiées de la base de prisme sont
- notées dans le sens trigonométrique à partir
d´une position temporale gauche - : 0°, 55°, 125°,
180°, 235°, 305°.
Tous les sujets de cette
étude ont réalisé un test de piétinement
sans prisme et un test de piétinement avec prisme dans
au moins une de ces positions de la base du prisme. Aucun sujet
n´a répété le test de piétinement
pour toutes les positions de la base du prisme.
La position de la base du
prisme étudiée sur un sujet donné a été
déterminée par les autres tests de son examen clinique
postural qui manifestaient la possibilité de modifier le
tonus postural de ce sujet par un prisme de trois dioptries prismatiques
placé dans cette position.
Résultats
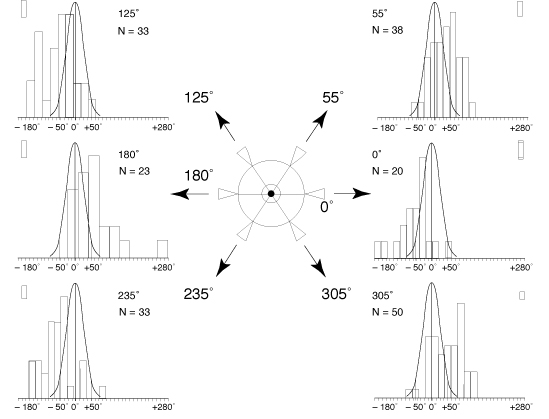
La distribution des variations
du mouvement de spin induite par le port du prisme est très
significativement différente de la distribution des variations
aléatoires de ce même mouvement (p<0,001), quelque soit la position de la base du prisme
(fig. 4).
Le sens de la variation du
mouvement de spin dépend de la position de la base du prisme
(fig. 4):
- tantôt, sous l´effet du prisme,
le sujet pivote davantage vers sa droite lorsque la base du prisme
est placée à 0° ou 125° ou 235° (valeurs
négatives de la différence des spins avec et sans
prisme),
- tantôt il pivote davantage vers sa
gauche lorsque la base du prisme est placée à 55°
ou 180° ou 305° (valeurs positives de la différence
des spins avec et sans prisme).
FIG. 4 - Variations du spin observées
selon les différentes positions de la base du prisme.
Au centre: diagramme présentant les différentes
positions étudiées de la base du prisme placé
devant l´oeil du patient.
Sur les côtés,
pour chacune des six positions de la base du prisme, histogramme
de la variation du mouvement de spin produit par la mise en place
du prisme. Courbe de Gauss rappelant la distribution normale théorique
des variations aléatoires du mouvement de spin.
Discussion
La portée de cette
analyse statistique est limitée. Elle ne concerne que des
sujets qui présentaient des troubles fonctionnels en rapport
avec la posture orthostatique. Une étude semblable cependant
a été faite sur des sujets normaux (Ushio et
al., 1980), mais elle ne concernait que les positions 0°
et 180° de la base du prisme et ne portait que sur un petit
nombre de sujets. Néanmoins les résultats de l´étude
d´Ushio et col. sont concordants avec la présente
analyse.
Chaque
sujet n´a été étudié que dans
une seule position de la base du prisme, les différences
décrites entre toutes les positions de la base du prisme
pourraient donc être rapportées à des différences
interindividuelles. Cependant une étude, proche des travaux
présentés dans cet article, a été
réalisée sur des sujets normaux avec des prismes
tournants, qui, en huit secondes, exploraient chez le même
sujet toutes les positions possibles de la base du prisme (Séverac
et al., 1993). Or les résultats de Séverac
et col. montrent aussi une différence de direction de la
réponse posturale selon la position de la base de prisme.
La concordance des résultats de Séverac et col.
avec les présents résultats s´arrête
là car les différences entre les protocoles ne permettent
pas de conclure plus précisément.
Conclusion
Les faits cliniques rapportés
ne permettent pas de prouver que la perception de l´espace,
au cours de l´ontogenèse, s´organise d´une
manière cohérente entre les canaux sensoriels vestibulaire
et visuel, mais ils rendent certainement cette hypothèse
encore plus probable.
Bibliographie
Adrian E.D. (1943) Discharges from
vestibular receptors in the cat. J. Physiol. (London),
101, 389-407.
Blakemore C, Van Sluyters RC, Peck CK, Hein A (1975) Development
of cat visual cortex following rotation of one eye. Nature,
257, 584-6
Blank R.H.I., Estes M.S., Markham C.H. (1975.a) Physiologic caracteristics
of vestibular first-order canal neurons in Cat. II. Response to
constant angular acceleration. J. Neurophysiol., 38,
1250-68.
Blanks R.H.I., Curthoys I.S., Markam C.H. (1975.b) Planar relationships
of the semicircular canals in man. Acta Otolaryngol. (Stockh.),
80, 191-200.
Cohen B., Suzuki J., Shanzer S., Bender M.B. (1964) Semi-circular
control of eye movements. In: M.B. Bender (Ed.) The oculomotor
system. Harper & Row, New York.
Cynader M, Berman N, Hein A (1975) Cats raised in a one-directional
world: effects on receptive fields in visual cortex and superior
colliculus. Exp Brain Res, 22, 267-80
Cynader M, Berman N, Hein A (1976) Recovery of function in cat
visual cortex following prolonged deprivation. Exp Brain Res,
25, 139-56
Estes M.S., Blanks R.H.I., Markham C.H. (1975) Physiologic caracteristics
of vestibular first-order canal neurons in Cat. I. Response plane
determination and resting discharge characteristics. J. Neurophysiol.,
38, 1232-49.
Fukuda T. (1959) The stepping test. Two phases of the labyrinthine
reflex. Acta Otolaryngol. (Stockh.) 50, 2: 95-108.
Gagey P.M. (1993) Le bilan postural. Ann. Kinésithér.,
20, 295-301.
Gagey P.M., Weber B. (1995) Posturologie; Régulation
et dérèglements de la station debout. Masson,
Paris.
Gagey P.M., Dujols A., Fouché B., Marucchi C. & Weber
B. (1994) The law of the canals: systematic variations of the
spin movement during Fukuda´s stepping test depends on the
position of the prism base. in: Vestibular and neural front.
Taguchi K., Igarashi M., Mori S. Elsevier, Amsterdam, 537-540.
Hein A (1987) La structuration de l´espace visuel peut-elle
se développer en l´absence des informations proprioceptives
oculomotrices? Critique de la Posturologie, 12,
1-4.
Hein A, Vital-Durand F, Salinger W, Diamond R (1979) Eye movements
initiate visual-motor development in the cat. Science,
204, 1321-2
Ito M., Nisimaru N., Yamamoto M. (1976) Pathways for the vestibulo-ocular
reflex excitation arising from semicircular canals of rabbits.
Exp. Brain Res., 24, 257-271.
Lorente de Nò R. (1932) The regulation of eye positions
and movements induced by labyrinth. Laryngoscope, 42,
233-332.
Löwenstein O. & Sand A. (1940) The individual and integrated
activity of the semicircular canals of the elasmobranch labyrinth.
J. Physiol. (London), 99, 89-101.
Magnus R. (1924) Körperstellung. Springer (Berlin).
Ross D.A. (1936) Electrical studies on the frog´s labyrinth.
J. Physiol. (London), 86, 117-46.
Séverac A., Bessou P., Pagès B. (1993) Unusual visual
stimulation in dynamic balance conditions: proposal for a space
motion sickness test. Advances in space research,
Szentágothaì J. (1950) The elementary vestibulo-ocular
reflex arc. J. Neurophysiol., 13, 395-407.
Ushio N., Hinoki M., Baron J.B., Gagey P.M., Meyer J. (1976) The
stepping test with neck torsion: proposal of a new equilibrium
test for cervical vertigo. Practica Otologica Kyoto, 69,
Sup.3, 1369-79 (En japonais).
Ushio N., Hinoki M., Nakanishi K., Baron J.B. (1980) Rôle
of oculomotor proprioception in the maintenance of body equilibrium;
correlation with the cervical one. Agressologie, 21,
E, 143-52.
Weber B., Gagey P.M., Noto R. (1984) La répétition
de l'épreuve modifie-t'elle l'exécution du test
de Fukuda? Agressologie, 25, 1311-14.
Wolgin DL, Hein A, Teitelbaum P (1980) Recovery of forelimb placing
after lateral hypothalamic lesions in the cat: parallels and contrasts
with development. J Comp Physiol Psychol, 94, 795-807
Zotterman Y. (1943) The microphonic effect of teleost labyrinth
and its biological significance. J. Physiol. (London),
102, 313-8.
Retour à l'accueil